
 This is a basic introduction to evolutionary biology and phylogenetic trees directed at undergraduate students (following the B. Sc. syllabus at the University of Delhi). It might also be of interest to high school level students (and lay persons) who want to learn something about this field.
This is a basic introduction to evolutionary biology and phylogenetic trees directed at undergraduate students (following the B. Sc. syllabus at the University of Delhi). It might also be of interest to high school level students (and lay persons) who want to learn something about this field.
After reading these pages, the reader should be able to:
- understand some basic principles of evolution
- understand ‘tree-thinking’
- the branching process of evolution and in what sense phylogenetic trees model this process
- read and interpret phylogenetic trees
- trace the evolution of characters on phylogenetic trees
- understand basics about the origin and evolution of angiosperms (flowering plants)
- understand basics about the coevolution of angiosperms and animals
Introduction
Evolution is descent with modification. Evolutionary biology is the study of both evolutionary history and evolutionary processes. Evolutionary history tells us what happened, when, and under what conditions. What, for instance, is the history of evolution of groups such as angiosperms, land plants, birds, or dinosaurs? When did they originate, diversify and go extinct? When, and in which groups, did particular traits, such as vascular tissue, evolve? Evolutionary processes are the factors that underlie evolutionary change. They help to answer questions such as, how did the close relationship between figs and fig wasps arise? What forces make populations change over time, make new species, and generate the huge range of biodiversity that exists on earth today?
Evolutionary Processes
The study of evolutionary processes tells us how populations and species evolve, i. e., how they change over time, speciate, and diversify. Such processes are most readily studied in populations or species (microevolution). These processes operate in all life systems and are major causes for differences we see within species as well as across species (macroevolution).
Microevolution
Species are made up of populations of individuals that can interbreed with one another, and these breeding populations are thought to be the smallest unit of evolution. The species (or, equivalently, the population) generally is thought to be the unit of evolution. Genetic variation among individuals within populations is the starting point of evolution. Variation may arise due to mutation and genetic recombination within individuals or migration of individuals (gene flow) in and out of populations. Mutation and migration are evolutionary forces that provide the basic “raw material” for evolution, but may not be strong enough on their own to change the traits of a population. For example, a mutation may arise in one or several individuals, but if it is not passed to the next generation, evolution cannot take place. On the other hand, natural selection and genetic drift are strong evolutionary forces that can lead to changes in the traits of a population, given variation. Population genetics is a field that tries to understand how genes behave in populations; these insights are used to predict changes in the frequency of a trait in a population under different conditions.
Imagine a population in which a gene governing the trait flower colour has a single form (allele A1) that results in white flowers. A mutation of this gene results in a new allele, A2, that results in red flowers. Birds in the area favour the red flowers; they carry pollen from one red flower to another, so plants bearing red flowers set more seeds than white-flowered plants (differential reproduction due to natural selection). Over several generations, continued natural selection on this trait results in the increase of red flowered plants (with allele A2) and decrease of white flowered plants (with allele A1) in the population. By the 200th generation, allele A2 increases to a frequency of 100% and is said to become fixed, while allele A1 goes down to a frequency of 0% and is said to become extinct.
In the example above, the red flower is an adaptation because plants that have red flowers are able to leave more offspring (are more fit) than plants that have white flowers (are less fit). Adaptations can only result from the evolutionary process of natural selection. Change could also occur due to the random survival and reproduction of individuals carrying a trait that is not under selection, by a process called genetic drift. Such a change does not result in adaptation. In other words, not all evolutionary change is adaptive.
Speciation
Many evolutionary changes over time may accumulate to such a point that a new species is said to have arisen.
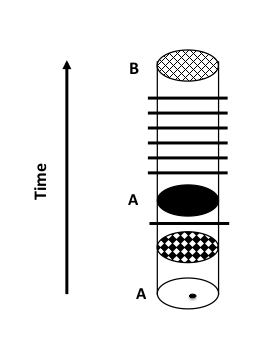
FIGURE 1a. Evolution through anagenesis. Xylosa anamika contains mostly white individuals; a black mutation arises in this white population and, over many generations, increases in frequency, until it eventually reaches 100% (fixation, denoted by the black horizontal bar). The population is still recognizable as Xylosa anamika. Over time, many other such changes accumulate (multiple bars) and the change in the population is so great that taxonomists now assign it to a new species, X. anonymosa.
This is the process of anagenesis, the formation of a new species within a lineage (Fig 1a). More commonly, new species are formed (i.e., speciation occurs) through the division of populations and species such that they no longer interbreed (reproductive isolation), and each newly isolated population evolves along its own lineage (Fig 1b). Over time, these isolated breeding populations will become new species, and the number of species increases due to repeated branching (cladogenesis). This is how the vast range of biological diversity on Earth was generated and is still being generated.
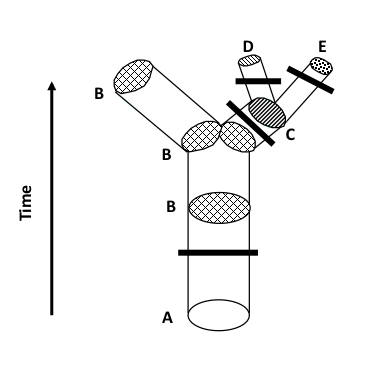
FIGURE 1b. Some time after X. anonymosa forms (multiple changes during anagenetic speciation represented by the thick bar), it is divided into two reproductively isolated populations that follow different evolutionary paths. One isolated population does not change much and continues to be recognized as Xylosa anonymosa, while the other undergoes many rapid changes to evolve into X. synonyma; this is cladogenetic speciation. X. synonyma, in turn, almost immediately speciates to form two new species, X. asymmetrica and X. symmetrica, again by cladogenetic speciation.
In cladogenetic speciation (1b), a key event is the division of an existing species into at least two reproductively isolated populations with no gene flow between them. Because they no longer form a single population with a single gene pool, they are effectively divided into independent populations that may now change due to the usual evolutionary processes. Over time, they may diverge from one another to the extent that they are recognizable as two distinct species. Thus, instead of the single ancestral species, there now are two descendant species.
Each of these species may itself undergo speciation events depending upon the play of evolutionary forces. A series of such speciation events represents a branching process, which may be visualized as a phylogenetic tree (Figure 2).
Macroevolution
Macroevolution occurs over geological time, or millions of years, and involves many species or other taxa (genera, families, classes, phyla), both extant and extinct. Macroevolution includes large scale patterns such as the history of change (evolution of the giraffe’s neck) and extinction of cycads and the dinosaurs that ate them. Macroevolutionary patterns and processes are revealed through the study of evolutionary history using methods of palaeontology and palaeobotany (geological and morphological study of fossils) as well as phylogenetic methods applied to morphological and molecular data of extant organisms. Evolutionary history includes understanding the origin and diversification of species and ‘higher’ taxa as well as the traits that they carry.
Phylogenetic Trees
A series of speciation events starting from a single ancestral species (Figure 1) represents a branching process (cladogenesis), so a branching phylogenetic tree (Figure 2) may be used to model the processes of evolution (microevolution, macroevolution, speciation and cladogenesis).
Reading Phylogenetic Trees
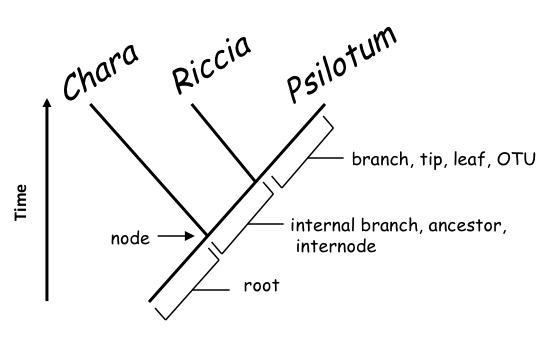
FIGURE 2. Figure 2. A rooted phylogenetic tree, showing relationships among three taxa Chara, Riccia and Psilotum. The tree consists of branches (representing species or lineages) and nodes (points where a branch splits, i.e., speciation events). Branches may be terminal (tip, leaf, OTU) or internal (ancestor, internode); a root is a special branch that serves to indicate the time axis: the root is in the past and the tips are in the present. In this fully resolved, dichotomous tree, Riccia and Psilotum are each others’ closest relatives (‘sisters’), while Chara is more distantly related to both.
A phylogenetic tree makes statements about (i) relationships among the taxa depicted, (ii) ancestors and descendants and (iii) speciation events in the history of the taxa. The tree consists of branches and nodes (Figure 2.) Each branch represents a species or a lineage; each node represents the point where a branch splits into two (speciation). Branches may be terminal (‘tips’ or Operational Taxonomic Units, OTU’s) or internal (actual or hypothetical ancestors). The root is a special branch that serves to indicate relative time in the tree. In a rooted tree, tips are more recent than the nodes from which they descended; nodes are more recent than the root.
A dichotomous tree (with only two descendant branches from a single ancestor, Figure 2) is fully resolved, because there is only one possible interpretation or resolution of the speciation event. A polychotomous tree may have one or more unresolved nodes. each with three or more branches descending from it (Figure 3 below).
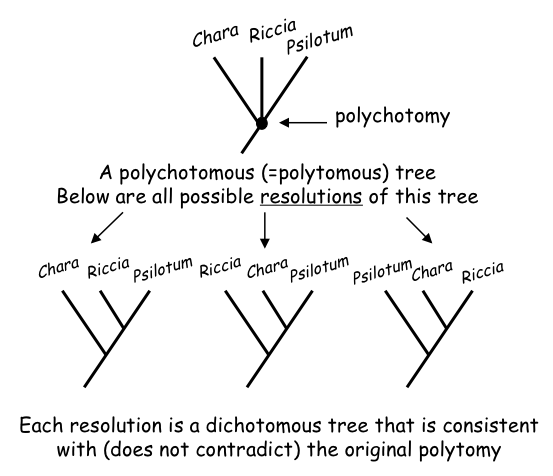
For each unresolved node there is more than one possible resolution of the order of the speciation events. Each resolution implies a distinct set of relationships among the species (Figure 3). Depending upon the study, unresolved nodes may represent either uncertainties in relationships (‘soft’ polytomy) or rapid speciation events (‘hard’ polytomy).

FIGURE 4. (a) In this cladogram of relationships among four taxa A, B, C, and D, (C, D) form a clade; (B, C, D) form a clade and (A, B, C, D) form a clade. C and D share a most recent common ancestor, and are therefore more closely related to each other than either is to A or B. The clade (C,D) is nested within the clade (B,C,D) which, in turn, is nested within the clade (A, B, C, D). C and D share a most recent common ancestor (3); B, C and D share a more recent common ancestor (2) than any of them do with A. Ancestor 1 gave rise to four descendants, A, B, C, D; ancestor 2 gave rise to three descendants, B, C, D; ancestor 3 gave rise to two descendants, C, D. (b) This cladogram shows that the relative positions of the taxa along the horizontal axis (along tips) is not relevant, but that the ancestor-descendant relationships are the same as in (a)
A monophyletic group (= clade) consists of a group of all the taxa that descended from an ancestral taxon (Figure 4). Clades are nested inside larger, more inclusive clades (‘higher’ taxa). In the graph represented by a phylogenetic tree, the axis along the tips has no scale or any significance: e.g., just the fact that two names are next to each other in a tree does not denote that they are each other’s closest relatives.
The tree is like a fixed mobile—swinging around a node does not alter relationships of taxa in the tree (Figure 5). In other words, it is important to read the phylogenetic tree from tips to nodes and not from tip to tip.

FIGURE 5. All these upside down trees represent the same relationships. Psilotum and Riccia are more closely related to each other than either is to Chara. This is true for all these tree ‘mobiles.’
Phylogenetic Tree, Cladogram, Phylogram
Evolutionary relationships among taxa are depicted as a branching diagram – a “tree” or “dendrogram.” Different terms are used for different types of dendrograms. In general, it is assumed that the evolutionary tree or phylogenetic tree depicts the evolutionary history of a lineage (= monophyletic group of taxa).
A cladogram (Fig 6a) shows relationships among entities, e.g., living taxa, extinct fossil taxa, sequences. It is a rooted tree that shows the closest relatives (sister taxa) descending from a most recent common ancestor. The root indicates the past and the tips indicate the present (assuming the OTUs are extant taxa). There is no scale associated with a cladogram — the lengths of branches do not signify anything. A cladogram may also display the history of particular traits.
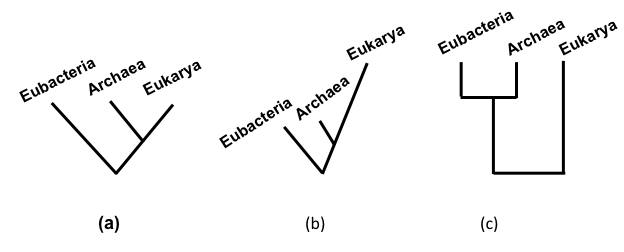
FIGURE 6. A dendrogram is any branching diagram or tree. (a) A cladogram is a dendrogram or phylogenetic tree that represents a hypothetical phylogeny. (b) A phylogram is phylogenetic tree in which branch lengths are proportional to the amount of inferred evolutionary change along the branches. (c) A phenogram is a dendrogram or tree that shows overall similarity among the terminal units.
Other evolutionary trees display information such as amount of evolutionary change (phylogram), extent of similarity (phenogram) or time of speciation events (chronogram).
A phylogram (Fig 6b) is a tree in which branch-lengths are drawn proportional to the amount of character evolution. Thus, a lineage represented by a long branch shows more change than one represented by a short branch. Branches of different lengths indicate that the rate of evolution of traits varies across different lineages. A phenogram (Fig 6c) is a rooted tree in which all tips are equally distant from the root. It is meant to show overall similarities among the OTUs. A phenogram can be used to represent evolutionary relationships only if there is no variation in the rate of evolution of traits, which is rarely the case.
These terms often are used carelessly and may have overlapping meanings depending on context and usage.
Why Clades?
We use phylogenetic trees to identify monophyletic groups or clades. A clade is a group of taxa that consists of a recent common ancestor and all of its descendants, so all taxa in this group have a shared history. Each taxon in a clade has its own unique history, but all of them also share a common evolutionary past because they descended from a common ancestor. Thus, monocots are a clade of diverse angiosperms that are more closely related to each other than to other angiosperms. The goal of phylogenetic systematics is to classify taxa according to their evolutionary history; the aim is to recognize, delimit and name only those groups of species that constitute clades.
Members of a polyphyletic group do not share a recent common ancestor, e.g., ‘algae.’ The group that has been named ‘algae’ consists of a group of taxa that are distantly related and scattered across the tree of life and therefore do not share a unique history (unlike, say, the flowering plants). A paraphyletic group consists of a common ancestor and some (but not all) of its descendants. Many groups that we are familiar with are paraphyletic, e.g., ‘dicotyledons.’
According to the current view of angiosperm relationships (Figure 7), monocots, rather than being a sister group to ‘dicots,’ are nested within a clade that contains some ‘dicots’ – Magnoliids and Eudicots.
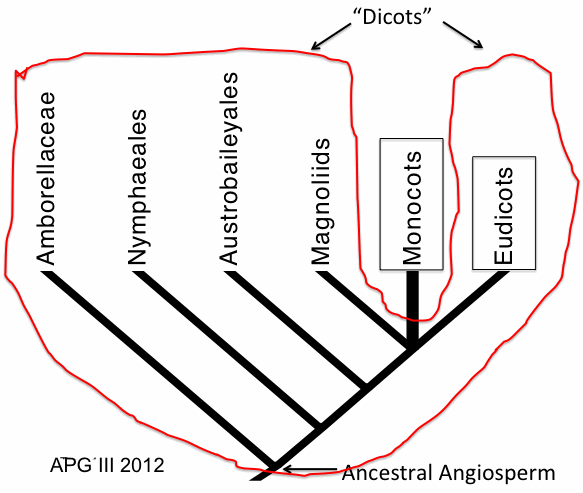
FIGURE 7. Phylogenetic Tree showing relationships among six major clades of angiosperms. Monocots are nested in a clade that contains Magnoliids and Eudicots. As a result, what used to be called dicots –Amborellaceae, Nymphaeales, Austrobaileyales, Magnoliids and Eudicots form a left-over paraphyletic (non-monophyletic) group. Following convention, this group is here labelled as “Dicots” to denote that it is a non-monophyletic, paraphyletic group.
As a result, the group that used to be called dicots and included Amborellaceae, Nymphaeales, Austrobaileyales, Magnoliids and Eudicots, is recognized to be a non-monophyletic (paraphyletic) group. The descendants of the most recent common ancestor of all ‘dicots’ also includes monocots, so by recognizing and naming the ‘dicots’ we are recognizing a paraphyletic group. In this paraphyletic group named ‘dicots’ not all descendants of an ancestor are included (i.e., monocots are excluded). Such a differentiation between monocots and ‘dicots’ ignores the fact that some ‘dicots’ (Magnoliids and Eudicots) share a more recent evolutionary history with monocots than with other ‘dicots’ (Amborellaceae, Nymphaeales and Austrobaileyales); this in turn may mean that these groups differ in some biological features. In other words, formal recognition and naming of the paraphyletic group ‘dicots’ misconstrues evolutionary history.
Evolution of Traits
Biologists are interested in understanding the history of not only organisms, but also that of the traits that these organisms carry. For instance, they might want to know about vascular tissue — when, and in which taxon did it evolve? Did it evolve just once in a common ancestor, so that the vascular tissue in these taxa is a homology? Did it evolve independently multiple times so that the presence of vascular tissue is due to homoplasy? The best way to understand this is by using a phylogenetic tree to reconstruct ancestral states across a tree and thus infer where, on the tree, transition from one state to another might have occurred.
For example, we take a well-supported phylogenetic tree and plot the traits at the tips (Figure 8a). One interpretation of the distribution of these data could be that vascular tissue evolved independently in each of these taxa; in other words, there were seven independent origins of the trait.
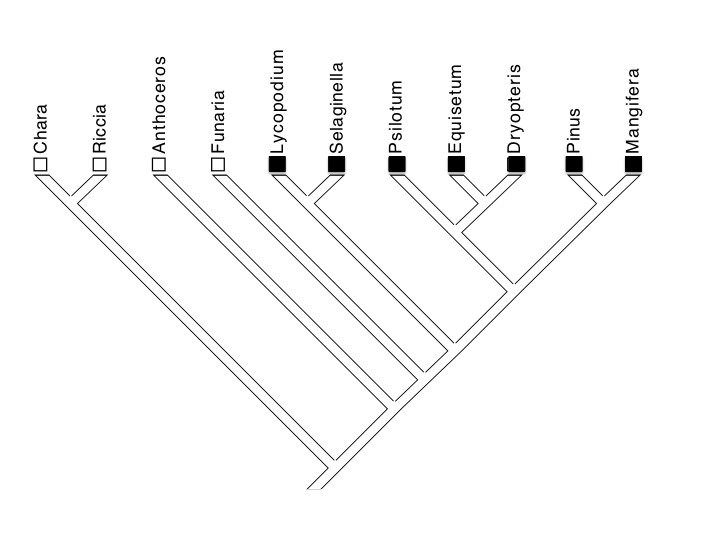
Fig 8a: Scenario 1 — Seven independent origins of vascular tissue in each of seven lineages of vascular plants
On the other hand, we may invoke the principle of maximum parsimony (hypothesize the fewest number of evolutionary changes or steps) to infer that there was a single origin of this trait in ancestor 5 (Fig 8b). Therefore, Pinus, Mangifera, Equisetum, Dryopteris, Psilotum, Lycopodium and Selaginella possess this trait because they inherited it from their most recent common ancestor, ancestor 5, which was a vascular plant. In other words, vascular tissue across the vascular plants is homologous because it arose once in this ancestral taxon.
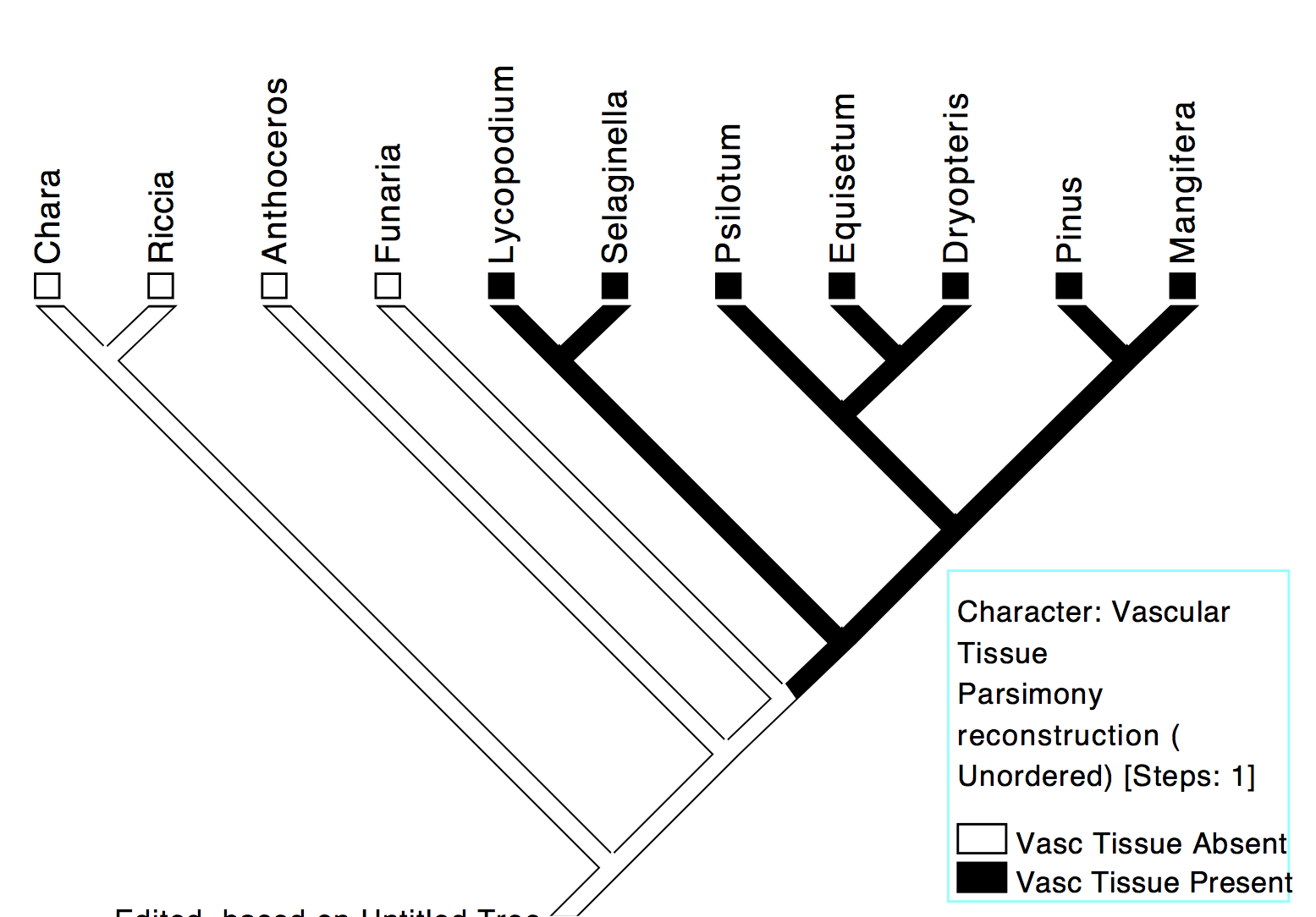
Fig 8b: Scenario 2 — A single origin of vascular tissue in the common ancestor (5) of vascular plants. The different vascular plants inherited their vascular tissue from this common ancestor, 5; in other words, vascular tissue in vascular plants is due to homology. This is the most parsimonious explanation of the taxonomic distribution of the trait.
Examining the evolution of vessels (specialized vascular cells), on the other hand, the most parsimonious inference is that there were three separate origins of vessels in Selaginella, Equisetum and Mangifera (Figure 8c).
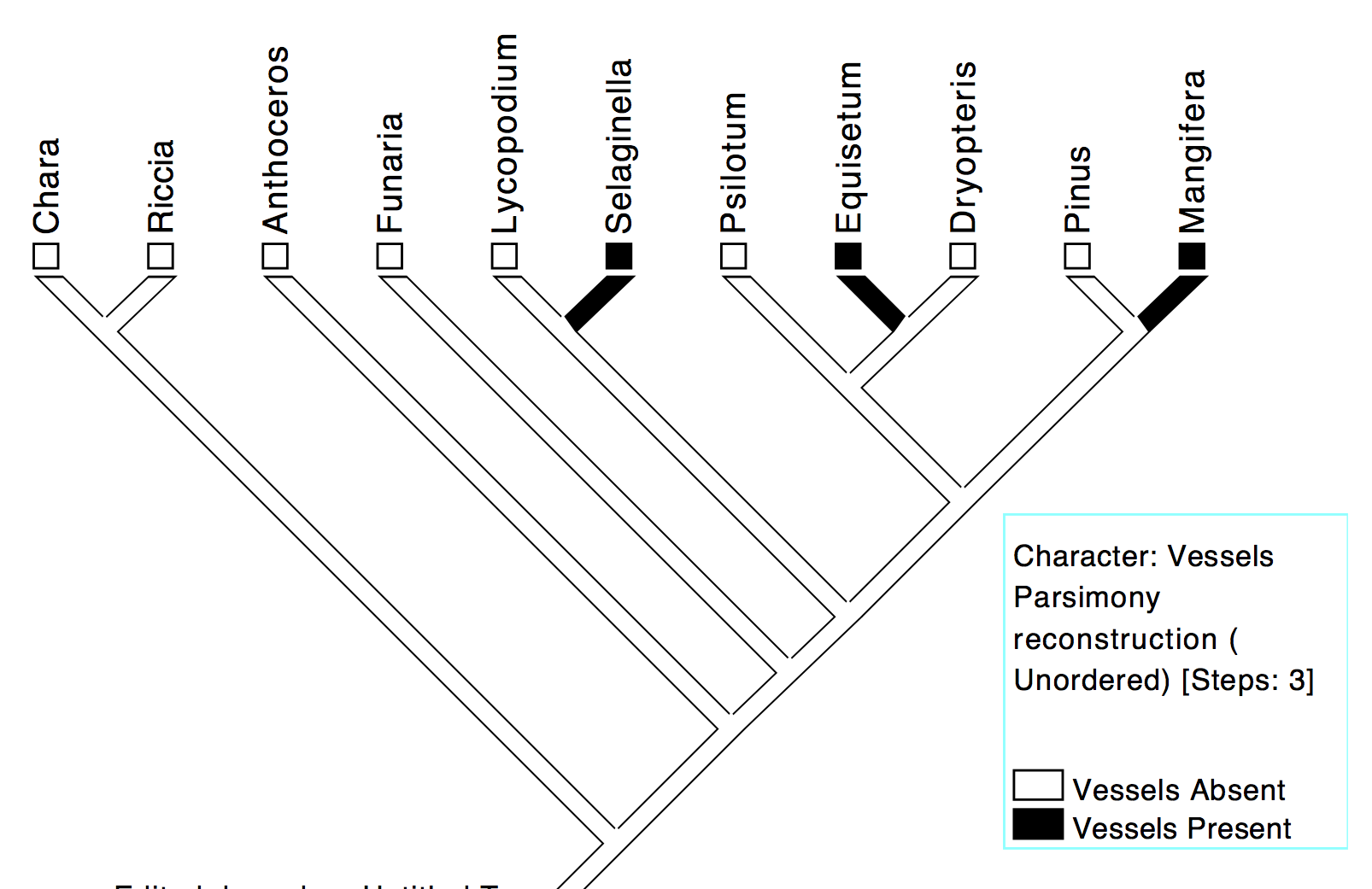
Fig 8c: Independent origins of vessels in three different lineages; the trait of vessels in these vascular plants is due to homoplasy (parallel evolution).
An alternative scenario could be that vessels evolved once in ancestor 5 and that independent losses occurred in Dryopteris, Psilotum, Lycopodium and Pinus, but this is less parsimonious–five steps (one gain and four losses) as against three gains–and therefore less acceptable. In other words, the presence of similar vascular cells called vessels in Mangifera, Equisetum and Selaginella is due to homoplasy, ie., independent parallel evolution of vessels in three distinct lineages.
Origin and Evolution of Angiosperms
“The abominable mystery” was how Darwin described the first appearance of many different types of angiosperms in the fossil record. This apparently sudden appearance suggested that either angiosperms had evolved for a long time in some hidden location/s or that they evolved very rapidly and produced diverse lineages within a short period. In either case, the pattern needs explanation.
Which are the closest relatives of angiosperms?
Angiosperms are characterized by a distinct combination of characters (tectate-columellate pollen, carpels, ovules with two integuments, highly reduced female gametophyte, double fertilization, endosperm, phloem consisting of sieve elements with companion cells), most of which cannot be preserved well, nor easily recognized, in fossils. Furthermore, the closest living relatives of angiosperms are difficult to identify using morphological data because, while double fertilization is found in some gymnosperms, other traits are not. Most recent molecular analyses suggest that gymnosperms are the closest living relatives of angiosperms.
The earliest cladistic analyses of seed plants suggested that angiosperms are part of the Anthophyte clade, which includes the flowering plants and fossils such as Pentoxylon (pdf), Glossopteris (pdf), Bennettitales and Caytonia — all of which have long been proposed as the closest relatives of angiosperms. This hypothesis is supported by recent phylogenetic analyses that combine molecular and morphological data.
Which were the early angiosperms?
There have been two different views on what flowers of the earliest angiosperms might have looked like. One view held that the flowers were unisexual with no perianth and male flowers producing large quantities of pollen, as in the present day walnut. According to the other view, early angiosperm flowers were large and cone like, with attractive perianth and large, spirally arranged stamens and carpels, much like present day Magnolia.
{kind=link}

Amborella trichopoda male flowers. By Scott Zona from USA [CC BY 2.0 (http://creativecommons.org/ licenses/by/2.0/deed.en)]

Nymphaea caerula By Leandro Avelar Brazil [CC BY-SA 4.0] https://creativecommons.org/licenses/by-sa/4.0/
and Nymphaeales (water lilies) with large bisexual flowers.

Archaefructus liaoningensis fossil. Shizhao – Own work. CC BY SA 2.5 [https://creativecommons.org/licenses/by-sa/2.5]
When did angiosperms originate and diversify?
The abominable mystery continues, even despite that large number of fossil angiosperms that are being unearthed across the world. The earliest angiosperms are known from the early Cretaceous, about 132.9 to 129.4 million years ago (Ma) in England, Israel and China. These determinations are based on pollen that are small, mono- or inaperturate and with reticulate-columellate walls. The earliest known angiosperm fossil showing the entire plant structure is Archaefructus, from about 125 Ma in China. The angiosperms, in general, show a pattern of very low diversity from first appearance (early Cretacous) to tremendous diversity within a relatively short period of time (late Cretaceous). This is particularly well documented in the Cretaceous deposits in eastern North America.
Co-evolution of Angiosperms and Animals
What is co-evolution? Paul Ehrlich and Peter Raven first enunciated the concept, by which two species are supposed to co-evolve if they undergo reciprocal evolution, i.e., evolutionary change in one species results in a corresponding change in the other and vice versa. Classic cases include defence responses of plants to their herbivores (e. g., butterflies) and plant-pollinator relationships such as those between figs and fig-wasps. Two types of co-evolution are considered: 1) evolutionary changes in the traits of one species eliciting corresponding evolutionary changes in the partner, and 2) co-speciation, where speciation in one species is accompanied by congruent speciation in the other partner. It should be noted that a more diffuse co-evolution between groups of species may also occur, and could be more common than a tight association between particular pairs of species.
Plant Defence and Herbivore Offence
Plant feeders (herbivores) have long been the bane of plant life, whether in the ocean, fresh-water bodies or on land. Structural (physical modification of surfaces or organs) as well as biochemical defences (including latex) against these voracious eaters are found widely in algal groups and green plants, including land plants. The herbivores against whom these defences are directed, of course, differ depending on the environment in which these interactions occur. The animals, in turn, evolve traits (‘offences’) that enable them to overcome defences put up by the plants they want to eat. Early vascular plants may have been eaten by biting and chewing herbivores. The fossil record suggests that, over time, defence-offence coevolution led to sap-sucking, gall-forming, leaf-mining and nectar-feeding herbivores (e.g., Labandeira et al. 1994).
Plant surfaces of angiosperms provide the first line of defence and may be covered by trichomes (hairs) that prevent insects from probing the tender tissue beneath. Thorns and spines provide protection against larger feeding animals such as goats and cattle. However, these physical defences may be overcome by animals that evolve the ability to deal with the obstacle. For instance, Passiflora lobata produces hooked trichomes that deter most insects, but the butterfly, Heliconia charithonia, has evolved to overcome this problem.
{kind=link}
{kind=link}
Dispersal
Animals, especially insects, birds and mammals, play an important role in the spread of reproductive propagules or structures of angiosperms. Dispersal of pollen ensures cross-breeding and the formation of a new generation, and dispersal of seeds ensures the establishment of a new generation in distant places. Dispersal spreads diverse genes among populations of species and expands the distribution of a species. Both pollen and seeds evolved in the seed plants, which include gymnosperms and angiosperms. However, most gymnosperms produce vast quantities of pollen, which is carried by the wind to ovules. Gymnosperm seeds, too, tend to be spread by wind (winged seeds) and rarely by birds (yew seeds with attractive red arils). Angiosperms have evolved the widest range of pollination mechanisms, which are related to the wide range of pollinators–insect, bird and mammal. Similarly, fruit dispersal mechanisms have depended on a large range of animals, including elephants and other large mammals now extinct. The huge diversity of angiosperm flowers and fruit has been thought to be the result of coevolution with their pollinators and seed dispersers.
Pollen
Insects, birds, and small mammals are the most common pollinators of flowers. Pollination syndromes are diverse assemblages of floral traits that evolve in response to selection by these varied pollinators. For example, bird-pollinated flowers often are relatively large, tubular, and produce large amounts of nectar. Floral traits may thus be used to predict associated pollinators. Some of these associations such as between figs and fig wasps and yucca and yucca moths exhibit strict co-evolution, as inferred from both intraspecific studies and phylogenetic analyses. In other cases, such as the South African Lapeirousia anceps (Iridaceae), reciprocal evolution of both flowers and pollinators is supported by studies of intraspecific variation.
Seed
In general, seed dispersers are attracted to fruit, which contain seeds, and the seeds are dispersed as a by-product of the consumption of fruit. Fruits adapted to attract dispersers may have bright colours and attractive smells; such fruits are fleshy, may be sweet or starchy, and the seeds generally are adapted to survive ingestion and defecation. The semi-parasitic mistletoe, Phoradendron (Viscaceae), grows in southwestern United States. Its seeds are especially dispersed by a bird, the silky flycatcher, that is, in turn, highly dependent on its fruit. This tight relationship is thought to be the result of coevolution, as both plant and bird have specific requirements and adaptations that make the interaction effective.
Summary
Evolutionary biology is the study of evolutionary processes and evolutionary history; it helps to explain the astonishing diversity of living organisms on earth. The evolutionary forces of mutation, migration, selective mating, natural selection and genetic drift contribute to genetic and phenotypic changes in successive generations of species. Most new species arise due to the splitting of existing species; such splitting leads to reproductive isolation of populations that, over the course of time (multiple generations) evolve into new species, Successive splitting of populations results in groups of species that that more closely related to each other than to other such groups of species. These groups are recognized by taxonomists (phylogenetic systematists) who identify, name and classify species into larger groups of ‘higher’ taxa. The monophyletic groups recognized in systematics are delimitied in phylogenetic trees, which are estimated using phylogenetic methods. Phylogenetic trees represent evolutionary history and are used to understand the origin and diversification of groups and their traits. The origin and history of angiosperms and their coevolution with animals is understood through a combination of methods including palaeobotany, systematics, phylogenetics and the study of evolutionary processes.
Further Reading
Futuyma, D. F. 2013. Evolution (Third edition. ed. ). Sunderland, Mass.: Sinauer. ISBN 978-1-60535-115-5.
Hall, B. K. and Hallgrimsson B. Strickberger’s Evolution. 2013. Jones and Bartlett, Sudbury, MA. SBN-13: 9781449691929
Zimmer, C. and Emlen D. J. 2013. Evolution: Making Sense of Life. Roberts & Co. ISBN 978-1-93622-117-2.
References
Labandeira C. C., Dilcher, D. L., Davis, D. R., and Wagner, D. L. 1994. Ninety-seven million years of angiosperm-insect association: paleobiological insights into the meaning of coevolution. PNAS 91: 12278–12282 [http://www.pnas.org/content/91/25/12278]
Acknowledgements
Thanks to Roshini Nayar, Beatrice Grabowski, Rama Shankaran and Usha Thorat for comments on previous versions.
Glossary
Adaptation (n.) A trait that occurs in a population because it gives an advantage to the organism carrying it. This advantage enables the organism to survive and reproduce better than individuals that do not carry the trait. The term adaptation (v.) has also been used to denote the process that leads to the emergence of adaptive traits, i.e., evolutionary change due to natural selection. The term adaptation (v.) is often (confusingly) used for acclimation, which is change in the phenotype of an individual organism in response to changed environmental conditions, i.e., which is developmental change.
Allele/s Alternative form/s (variations) of a gene at a particular location (locus) on a chromosome.
Anagenesis Evolutionary changes within a species or non-branching lineage leading to the formation of new species
Clade A monophyletic group. It consists of a recent common ancestor and all descendants of this ancestral taxon. Clades arise through a series of cladogenetic splitting (speciation) events.
Cladogenesis The evolutionary splitting of a species resulting in branching, and formation of two (or more) new species. A series of splitting events generates monophyletic groups of species (clades or branching lineages)
Evolutionary Forces Mechanisms that result in evolutionary change: mutation, migration, genetic drift and natural selection
Fitness The ability of a genotype to survive and reproduce and to bear offspring that carry on that genotype to the next generation. Flowers with sticky pollen may be more successful in pollination and therefore plants with genotypes that make such flowers are more successful in leaving offspring in the next generation. Fitness is relative – today a particular genotype (P1) may be more successful than another (P2), but a mutation in the future may yield genotype P3 which is even more successful than P1 and, therefore, will have a higher relative fitness than P1.
Fixation and extinction of alleles Fixation Extinction
Gene flow The movement of alleles among populations due to migration of individuals.
Gene Pool The entire set of alleles at different loci in a population. The gene pool is equivalent to gathering all gametes in a population and counting the numbers of different alleles of each gene locus.
Genetic Drift The evolutionary process by which allele frequencies fluctuate from one generation to another due to random sampling of reproductive individuals that will represent parents of the next generation. When the population size is large, these fluctuations are insigificant, but when the populaiton size is small, small fluctuations in numbers can have large effects over successive generations.
Hardy Weinberg Principle Constancy of allele and genotype frequencies from generation to generation in the absence of evolutionary forces; where genotype frequencies can be predicted from the allele frequencies.
Homology Similarity due to common ancestry. Two (or more) taxa have the same character state because they inherited the trait from a recent common ancestor that had the trait. The trait evolved in that common ancestor, and all its descendants inherited the trait.
Homoplasy Similarity due to parallelism, convergence or reversal. Two or more taxa have similar character states even though their common ancestor did not have the trait. The trait evolved independently in the taxa due to evolutionary factors acting separately in the different taxa..
Lineage Descendants of a single ancestor. The lineage may be non-branching (a successive series of populations of a species descendant from a single ancestral species) or branching (a monophyletic group of species descendant from a single ancestral species).
Migration The movement of individuals, seeds, reproductive structures such as pollen, from one population to another, resulting in gene (allele) flow.
Monophyletic group A clade; a group consisting of an ancestral taxon and all its descendants.
Mutation Change in a gene that is random with respect to the environment of an organism
Natural Selection The differential survival and reproduction of individuals in a population due to the fact that they carry a trait that gives them some kind of advantage over other individuals. Due to this advantage, these individuals are able to survive or reproduce better, leaving more offspring in the next generation. If the advantageous trait is heritable, then that trait is passed on to the next generation and the frequency of the trait increases over time.
Non-monophyletic group A group of species that do not include all descendents of their most recent common ancestor. Such a group may be termed paraphyletic when the component taxa share a recent common ancestor, but one or more clades are excluded from the larger group, e.g., ‘dicotyledons,’ which exlude monocotyledons. In a polyphyletic group component taxa do not share a recent common ancestor, e.g., ‘algae.’
Population A group of interbreeding individuals of a species that occur in a geographic region at the same time.
Population genetics The study of changes in frequencies of genes and alleles in a population and the connection of such changes with evolution.
Reproductive Isolation The cessation of sexual reproduction among subgroups of a population due to a range of abiotic and biotic factors such as geographic isolation, polylpoidy etc.
Rooting. A rooted tree shows the common ancestor of all the OTUs or taxa in that tree. This provides a way to show relative time going from the past, as shown by the root, and the present, as shown by the tips; this allows the narration of evolutionary history. An unrooted does not assign a single ancestral root and is instead compatible with several rooted trees (but not all possible trees). Rooting is most often done using the outgroup/s in the tree.
Selection
Speciation The process of formation of new species through either anagenesis or cladogenesis.
Taxon (Taxon s., Taxa pl.) Any named group or unit in a biological system of classification. In the phylogenetic system of classification, a taxon must be composed of all descendants of a single ancestor (i.e., represent a monophyletic group)
Trait Any feature or property (usually phenotypic) of an organism.
==========================================
This work is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License.